ما هي بنية ووظائف مجمع جولجي. جهاز جولجي
جهاز جولجي عبارة عن كومة من الحويصلات الغشائية المسطحة ("") ونظام الحويصلات المرتبطة بها. عند دراسة الأقسام شديدة الرقة ، كان من الصعب الكشف عن هيكلها ثلاثي الأبعاد ، ومع ذلك ، اقترح العلماء أن الأنابيب المترابطة قد تشكلت حول الجزء المركزي.
يقوم جهاز جولجي بوظيفة نقل المواد والتعديل الكيميائي للمنتجات الخلوية التي تدخله. هذه الوظيفة مهمة بشكل خاص في الخلايا الإفرازية ، على سبيل المثال ، تفرز خلايا أسينار البنكرياس الإنزيمات الهضمية لعصير البنكرياس في القناة الإخراجية. درس العلماء أداء جهاز جولجي باستخدام صورة مجهرية إلكترونية لمثل هذه الخلية. تم تحديد مواد النقل الفردية باستخدام الأحماض الأمينية ذات العلامات الإشعاعية.
تُبنى البروتينات من الأحماض الأمينية في الخلية. ثبت أنها تتركز في حويصلات جهاز جولجي ثم تنتقل إلى غشاء البلازما. في المرحلة النهائية ، يتم إفراز الإنزيمات غير النشطة ، ويكون شكل مماثل ضروريًا حتى لا يتمكنوا من تدمير الخلايا التي تتكون فيها. كقاعدة عامة ، البروتينات التي تدخل مجمع جولجي هي بروتينات سكرية. هناك يخضعون لتعديل يحولهم إلى علامات تسمح لك بتوجيه البروتين بدقة للغرض المقصود منه. لم يتم تحديد كيفية توزيع مجمع جولجي للجزيئات بدقة.
وظيفة إفراز الكربوهيدرات
في بعض الحالات ، يشارك جهاز جولجي في إفراز الكربوهيدرات ، على سبيل المثال ، في النباتات ، في تكوين مادة جدار الخلية. تم تعزيز نشاطها في منطقة لوحة الخلية ، الواقعة بين نواتين ابنتيتين حديثتي التكوين. يتم توجيه حويصلات جولجي إلى هذا الموقع بواسطة الأنابيب الدقيقة. تصبح أغشية الحويصلة جزءًا من أغشية البلازما للخلايا الوليدة. يصبح محتواها ضروريًا لبناء الجدران الخلوية للصفيحة المتوسطة والجدران الجديدة. يتم توفير السليلوز للخلايا بشكل منفصل عن طريق الأنابيب الدقيقة التي تتجاوز جهاز جولجي.
يقوم جهاز جولجي أيضًا بتصنيع مخاط البروتين السكري ، والذي يشكل المخاط في المحلول. يتم إنتاجه عن طريق الخلايا الكأسية ، التي تقع في سمك ظهارة الغشاء المخاطي في الجهاز التنفسي وبطانة الأمعاء. في بعض النباتات الحشرية ، في غدد الأوراق ، ينتج جهاز جولجي الإنزيمات والمخاط اللزج. يشارك مجمع جولجي أيضًا في إفراز الشمع والمخاط والعلكة والغراء النباتي.
جهاز جولجي
تشكل الشبكة الإندوبلازمية وغشاء البلازما وجهاز جولجي نظامًا لغشاء خلية واحدة ، حيث تحدث عمليات تبادل البروتين والدهون بمساعدة نقل الغشاء داخل الخلايا الموجه والمنظم.
تتميز كل عضيات غشائية بتكوين فريد من البروتينات والدهون.
هيكل AG
يتكون AG من مجموعة من الأكياس الغشائية المسطحة - صهاريج، مجمعة في أكوام - الديكتوسومات(حوالي 5-10 صهاريج ، في حقيقيات النوى السفلية> 30). يختلف عدد الديكتوسومات في الخلايا المختلفة من 1 إلى 500.
صهاريج منفصلة للديكتوسوم بسمك متغير - في وسط أغشيته يتم تجميعها معًا - يبلغ التجويف 25 نانومتر ، وتتكون الامتدادات على الأطراف - أمبولاتالذي عرضه غير ثابت. حوالي 50 نانومتر -1 ميكرومتر فقاعات من الأمبولات المتصلة بالصهاريج بواسطة شبكة من الأنابيب.
في الكائنات متعددة الخلايا ، يتكون AG من أكوام من الصهاريج مترابطة في نظام غشاء واحد. AG هو نصف كرة ، قاعدته تواجه النواة. يتم تمثيل الخميرة AG بواسطة صهاريج مفردة معزولة محاطة بحويصلات صغيرة وشبكة أنبوبية وحويصلات إفرازية وحبيبات. طفرات الخميرة Sec7 و Sec14 لها بنية تشبه كومة من صهاريج خلايا الثدييات.
تتميز AG بقطبية هياكلها. كل كومة لها قطبان: القطب القريب(تشكيل ، سطح رابطة الدول المستقلة) و القاصي(ناضجة ،
عبر السطح). قطب رابطة الدول المستقلةجانب الغشاء الذي تندمج به الفقاعات. عبر القطبجانب الغشاء الذي تنطلق منه الحويصلات.
خمس حجرات وظيفية من AG:
1. الهياكل الأنبوبية الحويصليّة الوسيطة (VTC أو ERGIC - الحيز الوسيط ER-Golgi)
2. Cis-cistern (cis) - الخزانات الموجودة بالقرب من ER:
3. الخزانات المتوسطة (الوسطية) - الخزانات المركزية
4. الصهريج العابر (العابر) - الأبعد عن خزانات الطوارئ.
5. الشبكة الأنبوبية المجاورة للمحول - شبكة جولجي العابرة (TGN)
تكون أكوام الصهاريج منحنية بحيث تواجه الطبقة الخارجية المقعرة القلب.
في المتوسط ، هناك 3-8 صهاريج في AG ، وفي خلايا الإفراز النشط قد يكون هناك المزيد (حتى 13 في الخلايا الخارجية للبنكرياس).
يحتوي كل خزان على أسطح رابطة الدول المستقلة وعابرة. تدخل البروتينات المركبة ، والدهون الغشائية ، والجليكوزيلاتي في ER ، AG من خلال القطب cis. المواد من خلال أكوام يتم نقلها عن طريق النقل
حويصلات منفصلة عن الأمبولات. عندما تمر البروتينات أو الدهون عبر مكدسات Golgi ، فإنها تخضع لسلسلة من التعديلات اللاحقة للترجمة ، بما في ذلك التغييرات في السكريات القليلة المرتبطة بـ N:
رابطة الدول المستقلة: Mannosidase I قلص سلاسل المانوز الطويلة إلى M-5
متوسط: N-acetylglucosamine transferase I ينقل N-acetylglucosamine
نشوة: السكريات النهائية المضافة - بقايا الجالاكتوز وحمض السياليك.
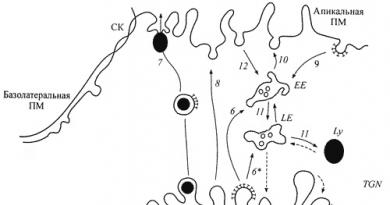
هيكل جهاز جولجي ومخطط النقل. خمسة مكونات AG ومخطط النقل:وسيطة (ERGIC) ، رابطة الدول المستقلة ، وسيطة ، وعابرة وعابرة لشبكة جولجي (TGN). 1. دخول البروتينات المركبة ، والبروتينات السكرية الغشائية ، والإنزيمات الليزوزومية في خزان ER العابر المجاور لـ AG ، و 2 - خروجها من ER في حويصلات تحدها COPI (النقل المتقدم). 3 - إمكانية نقل البضائع من الأنبوب الحويصلي
خمسة مكونات AG ومخطط النقل:وسيطة (ERGIC) ، رابطة الدول المستقلة ، وسيطة ، وعابرة وعابرة لشبكة جولجي (TGN). 1. دخول البروتينات المركبة ، والبروتينات السكرية الغشائية ، والإنزيمات الليزوزومية في خزان ER العابر المجاور لـ AG ، و 2 - خروجها من ER في حويصلات تحدها COPI (النقل المتقدم). 3 - إمكانية نقل البضائع من الأنبوب الحويصلي
مجموعات إلى خزان الصهريج لـ AG في حويصلات COPI ؛ 3 * - نقل البضائع من الخزانات السابقة إلى الخزانات اللاحقة ؛ 4 - إمكانية النقل الحويصلي الرجعي للبضائع بين صهاريج AG ؛ 5 - عودة البروتينات المقيمة من AG إلى tER باستخدام حويصلات تحدها COPI (النقل الرجعي) ؛ 6 و 6 * - نقل الإنزيمات الليزوزومية عن طريق الحويصلات المطلية بالكالاثرين ، على التوالي ، إلى EE المبكر و LE endosomes المتأخرة ؛ 7 - إفراز منظم للحبيبات الإفرازية ؛ 8 - التضمين التأسيسي لبروتينات الغشاء في غشاء البلازما القمي للـ PM ؛ 9 ، الالتقام الخلوي بوساطة مستقبلات مع حويصلات مغلفة بالكالاثرين ؛ 10 عودة عدد من المستقبلات من الإندوسومات المبكرة إلى غشاء البلازما ؛ 11 - نقل الروابط من EE إلى LE و Ly lysosomes ؛ 12- نقل الربيطات في حويصلات غير كلاذرين.
وظائف AG
1. المواصلات- ثلاث مجموعات من البروتينات تمر عبر AG: بروتينات الغشاء المحيط بالبروتينات ، والبروتينات المخصصة لها
للتصدير من الخلية والإنزيمات الليزوزومية.
2. فرزللنقل: الفرز من أجل النقل الأطول إلى العضيات ، PM ، الإندوسومات ، تحدث الحويصلات الإفرازية في مجمع جولجي العابر.
3. إفراز- إفراز المنتجات المصنعة في الخلية.
3. الارتباط بالجليكوزيلالبروتينات والدهون: جليكوزيداتإزالة بقايا السكر - إزالة الجليكوزيل ، جليكوزيل ترانسفيرازاتيعلقون السكريات مرة أخرى إلى سلسلة الكربوهيدرات الرئيسية - الارتباط بالجليكوزيل.يحدث فيه الارتباط بالجليكوزيل في سلاسل قليلة السكريد من البروتينات والدهون ، وكبريتات عدد من السكريات وبقايا التيروزين للبروتينات ، بالإضافة إلى تنشيط سلائف هرمونات البولي ببتيد والببتيدات العصبية.
4. توليف السكريات- يتم تكوين العديد من السكريات في AG ، بما في ذلك البكتين والهيميسليلوز ، والتي تشكل جدران الخلايا للنباتات ومعظم الجليكوزامينوجليكان ، والتي تشكل المصفوفة خارج الخلية في الحيوانات
5. الكبريت- معظم السكريات المضافة إلى لب البروتين في البروتيوغليكان تكون كبريتات
6. إضافة مانوز 6 فوسفات: يضاف M-6-P لتوجيه الإشارة إلى الإنزيمات المخصصة للجسيمات الحالة.
جليكوسيل
تبدأ معظم البروتينات في أن تصبح غليكوزيلاتي في ER الخام عن طريق إضافة السكريات القليلة المرتبطة بـ N إلى سلسلة البولي ببتيد المتنامية. إذا كان البروتين السكري مطويًا في الشكل المطلوب ، فإنه يخرج من ER وينتقل إلى AG ، حيث يخضع لتعديل ما بعد الترجمة.
تشارك إنزيمات Glycosyltransferase في الارتباط بالجليكوزيل للمنتجات المفرزة. إنهم يشاركون في إعادة تشكيل السلاسل الجانبية قليلة السكاريد المرتبطة بـ T وإضافة أجزاء السكاريد المرتبطة بـ O وأجزاء قليلة السكاريد من بروتين غليكوليبيد. .
بالإضافة إلى ذلك ، يحدث الارتباط بالجليكوزيل في مجالات غشاء البروتين الدهني ، والتي تسمى الأطواف ، في AG.
دولي كولفوسفاتيضيف مركب كربوهيدرات - 2GlcNAc-9-mannose-3-glucose إلى أسباراجين البولي ببتيد المتنامي. ينقسم الجلوكوز الطرفي إلى خطوتين: جلوكوزيداز أنايشق من بقايا الجلوكوز الطرفية ، الجلوكوزيداز الثانييزيل اثنين من بقايا الجلوكوز. ثم يتم قطع المانوز. في هذه المرحلة ، تكتمل المرحلة الأولية من معالجة الكربوهيدرات في ER وتدخل البروتينات التي تحمل مركب oligosaccharide إلى AG.
تتم إزالة ثلاث بقايا مانوز أخرى في خزانات AG الأولى. في هذه المرحلة ، يحتوي المجمع الأساسي على 5 مخلفات مانوز أخرى. N-acetylglucosamine transferase أنايضيف بقايا N-acetylglucosamine GlcNAc. يتم قطع ثلاث بقايا مانوز أخرى من المجمع الناتج. يتكون الآن من جزيئين GlcNAc-3-mannose-1-GlcNAc وهو بنية أساسية تضيف إليها ناقلات الجليكوزيل جزيئات أخرى
الكربوهيدرات. يتعرف كل ناقلة جليكوزيل على بنية الكربوهيدرات النامية ويضيف السكاريد الخاص به إلى السلسلة.
إفراز
مخطط الإفراز:
تتركز البروتينات التي تم تصنيعها في ER في مواقع الخروج من ER الانتقالية بسبب نشاط مجمع قسيم الغلاف COPII والمكونات ذات الصلة ويتم نقلها إلى حجرة ERGIC الوسيطة بين ER و AG ، والتي تنتقل منها إلى AG في مهدها حويصلات أو على طول الهياكل الأنبوبية. يتم تعديل البروتينات تساهميًا أثناء مرورها عبر خزانات AG ، ويتم فرزها على السطح العابر لـ AG ، وإرسالها إلى وجهاتها. يتطلب إفراز البروتين الدمج السلبي لمكونات غشاء جديدة في غشاء البلازما. يتم استعادة توازن الغشاء عن طريق الالتقام المستمر بوساطة المستقبلات.
المسارات الداخلية والخارجية للنقل الغشائي لها أنماط مشتركة في اتجاه حركة ناقلات الغشاء إلى ما يقابلها
الهدف وخصوصية الاندماج والتبرعم. نقطة الالتقاء الرئيسية لهذه المسارات هي AG.
مجمع جولجيعبارة عن كومة من الأكياس الغشائية (الخزان) ونظام من الفقاعات المرتبطة بها.
على الجانب الخارجي المقعر من كومة الحويصلات ، تتبرعم من الملساء. يتم تشكيل صهاريج جديدة EPS باستمرار ، وفي داخل الصهاريج تتحول مرة أخرى إلى فقاعات.
تتمثل الوظيفة الرئيسية لمركب جولجي في نقل المواد إلى السيتوبلازم والبيئة خارج الخلية ، بالإضافة إلى تخليق الدهون والكربوهيدرات. يشارك مجمع جولجي في نمو وتجديد غشاء البلازما وفي تكوين الجسيمات الحالة.
تم اكتشاف مجمع جولجي في عام 1898 من قبل ك.غولجي. بمعدات بدائية للغاية ومجموعة محدودة من الكواشف ، توصل إلى اكتشاف ، بفضله حصل مع Ramon y Cajal على جائزة نوبل. عالج الخلايا العصبية بمحلول ثنائي كرومات ، وبعد ذلك أضاف نترات الفضة والأوزميوم. بمساعدة ترسيب الأوزميوم أو الأملاح الفضية مع الهياكل الخلوية ، اكتشف جولجي شبكة داكنة اللون في الخلايا العصبية ، والتي أطلق عليها اسم الجهاز الشبكي الداخلي. عندما تكون ملطخة بالطرق العامة ، فإن المركب الرقائقي لا يتراكم الأصباغ ؛ لذلك ، تكون منطقة تركيزه مرئية كمنطقة ضوئية. على سبيل المثال ، بالقرب من نواة خلية البلازما ، تكون منطقة الضوء مرئية ، المقابلة للمنطقة التي توجد بها العضية.
غالبًا ما يكون مجمع جولجي مجاورًا للنواة. تحت المجهر الضوئي ، يمكن توزيعها في شكل شبكات معقدة أو مناطق منفصلة منتشرة (ديكتوسومات). لا يشكل شكل وموقع العضية أهمية أساسية وقد يتغير اعتمادًا على الحالة الوظيفية للخلية.
مجمع جولجي هو مكان تكاثف وتراكم نواتج الإفراز المنتجة في أجزاء أخرى من الخلية ، وبشكل رئيسي في EPS. أثناء تخليق البروتين ، تتراكم الأحماض الأمينية ذات النظائر المشعة في غرام. EPS ، ومن ثم تم العثور عليها في مجمع جولجي ، أو الشوائب الإفرازية أو الجسيمات الحالة. هذه الظاهرة تجعل من الممكن تحديد أهمية مجمع جولجي في العمليات التركيبية في الخلية.
يُظهر الفحص المجهري الإلكتروني أن مجمع جولجي يتكون من مجموعات من الصهاريج المسطحة التي تسمى الديكتوسومات. الدبابات متاخمة لبعضها البعض على مسافة 20 ... 25 نانومتر. يبلغ تجويف الخزانات في الجزء المركزي حوالي 25 نانومتر ، ويتم تشكيل امتدادات على الأطراف - أمبولات ، لا يكون عرضها ثابتًا. يوجد حوالي 5… 10 خزانات في كل كومة. بالإضافة إلى الصهاريج المسطحة المتباعدة بكثافة ، يوجد عدد كبير من الحويصلات الصغيرة في منطقة مجمع جولجي ، خاصة على طول حواف العضية. في بعض الأحيان يتم ربطهم من الأمبولات.
على الجانب المجاور لـ ER والنواة ، يحتوي مجمع Golgi على منطقة تحتوي على عدد كبير من الحويصلات الصغيرة والصهاريج الصغيرة.
مجمع جولجي مستقطب ، أي غير متجانس نوعيًا من زوايا مختلفة. لها سطح رابطة الدول المستقلة غير ناضج أقرب إلى النواة وسطح متحولة ناضج يواجه سطح الخلية. وفقًا لذلك ، تتكون العضية من عدة حجرات مترابطة تؤدي وظائف محددة.
تواجه حجرة رابطة الدول المستقلة عادةً مركز الخلية. سطحه الخارجي محدب الشكل. تندمج الحويصلات المجهرية (نقل الحويصلات الدبوسية) ، متجهة من EPS ، مع الصهاريج. يتم تجديد الأغشية باستمرار عن طريق الحويصلات ، والتي بدورها تعمل على تجديد محتويات التكوينات الغشائية للمقصورات الأخرى. تبدأ المعالجة اللاحقة للبروتينات في الحجرة وتستمر في الأجزاء التالية من المجمع.
تقوم المقصورة الوسيطة بإجراء عملية الارتباط بالجليكوزيل والفسفرة والكربوكسيل وكبريتات معقدات بروتين البوليمر الحيوي. يحدث ما يسمى بالتعديل اللاحق للترجمة لسلاسل البولي ببتيد. هناك توليف من الجليكوليبيدات والبروتينات الدهنية. في المقصورة المتوسطة ، كما هو الحال في حجرة رابطة الدول المستقلة ، تتشكل مجمعات البروتين من الدرجة الثالثة والرباعية. تخضع بعض البروتينات لعملية تحلل جزئي للبروتين (تدمير) ، مصحوبًا بتحولها الضروري للنضج. وبالتالي ، فإن المقصورات الوسيطة و cis مطلوبة لنضج البروتينات ومركبات البوليمر الحيوي المعقدة الأخرى.
تقع المقصورة العابرة بالقرب من محيط الخلية. سطحه الخارجي مقعر عادة. جزئيًا ، يمر الحيز العابر إلى الشبكة العابرة - وهو نظام من الحويصلات والفجوات والأنابيب.
في الخلايا ، يمكن توصيل الديكتوسومات الفردية ببعضها البعض عن طريق نظام من الحويصلات والصهاريج المجاورة للنهاية البعيدة لمجموعة من الأكياس المسطحة ، بحيث يتم تشكيل شبكة ثلاثية الأبعاد فضفاضة ، الشبكة العابرة.
في هياكل المقصورة العابرة والشبكة ، يحدث فرز البروتينات والمواد الأخرى ، وتشكيل حبيبات إفرازية ، وسلائف الجسيمات الأولية ، وحويصلات إفراز عفوية. الحويصلات الإفرازية و prelysosomes محاطة بالبروتينات - clathrins.
تترسب مادة الكلاذرين على غشاء الحويصلة الناشئة ، مما يؤدي إلى فصلها تدريجيًا عن الصهريج البعيد للمجمع. تخرج الحويصلات ذات الحدود من الشبكة العابرة ، وتعتمد حركتها على الهرمونات وتتحكم فيها الحالة الوظيفية للخلية. تتأثر عملية نقل الحويصلات المجاورة بالأنابيب الدقيقة. تتفكك مجمعات البروتين (الكلاذرين) حول الحويصلات بعد انشقاق الحويصلة من الشبكة العابرة وإعادة تشكيلها في لحظة الإفراز. في لحظة الإفراز ، تتفاعل مجمعات بروتين الحويصلة مع بروتينات الأنابيب الدقيقة ، ويتم نقل الحويصلة إلى الغشاء الخارجي. لا تُحاط حويصلات الإفراز العفوي بالكلاذرين ، بل يحدث تكوينها باستمرار وتتجه نحو غشاء الخلية ، وتندمج معه ، مما يضمن استعادة السيتوليما.
بشكل عام ، يشارك مجمع جولجي في الفصل - وهذا هو الفصل ، وفصل أجزاء معينة من الكتلة الرئيسية ، وتراكم المنتجات المصنعة في EPS ، في إعادة ترتيبها الكيميائي ، نضجها. في الخزانات ، يتم تصنيع السكريات ، ويتم دمجها مع البروتينات ، مما يؤدي إلى تكوين مجمعات معقدة من الببتيدوغليكان (البروتينات السكرية). بمساعدة عناصر مجمع جولجي ، تتم إزالة الأسرار الجاهزة خارج الخلية الإفرازية.
تنفصل فقاعات النقل الصغيرة عن الجرام. EPS في مناطق خالية من الريبوسومات. تقوم الفقاعات باستعادة أغشية مجمع جولجي وتوصيل معقدات البوليمر المركبة في EPS. يتم نقل الحويصلات إلى حجرة رابطة الدول المستقلة حيث تلتحم بأغشيتها. وبالتالي ، يتلقى مجمع جولجي أجزاء جديدة من الأغشية والمنتجات المصنعة في غرام. EPS.
في خزانات مجمع جولجي ، تحدث تغييرات ثانوية في البروتينات المركبة في غرام. EPS. ترتبط هذه التغييرات بإعادة ترتيب سلاسل قليل السكاريد للبروتينات السكرية. داخل تجاويف مجمع جولجي ، يتم تعديل البروتينات الليزوزومية وبروتينات الإفراز بمساعدة ترانسجلوكوزيدات: هناك استبدال ونمو متسلسلان لسلاسل قليل السكاريد. تنتقل البروتينات المعدلة من صهريج حجرة رابطة الدول المستقلة إلى صهاريج المقصورة العابرة عن طريق النقل في الحويصلات المحتوية على البروتين.
في المقصورة العابرة ، يتم فرز البروتينات: توجد مستقبلات البروتين على الأسطح الداخلية لأغشية الخزانات ، والتي تتعرف على البروتينات الإفرازية وبروتينات الغشاء والليزوزومات (الإنزيمات المائية). نتيجة لذلك ، تنفصل ثلاثة أنواع من الفجوات الصغيرة عن المواقع العابرة البعيدة للديكتوزومات: prelysosomes التي تحتوي على hydrolases ؛ مع شوائب إفرازية ، فجوات تجدد غشاء الخلية.
تتمثل الوظيفة الإفرازية لمركب جولجي في أن البروتين المصدر المركب على الريبوسومات ، والذي يتم فصله وتجميعه داخل خزانات EPS ، يتم نقله إلى فجوات الجهاز الرقائقي. ثم يمكن أن يتكثف البروتين المتراكم ، مكونًا حبيبات بروتينية إفرازية (في البنكرياس والثدي والغدد الأخرى) ، أو يبقى في شكل مذاب (الغلوبولين المناعي في خلايا البلازما). تنفصل الحويصلات التي تحتوي على هذه البروتينات عن الامتدادات الأمبولية لخزانات مجمع جولجي. يمكن أن تندمج هذه الحويصلات مع بعضها البعض ، وتزيد في الحجم ، وتشكل حبيبات إفرازية.
بعد ذلك ، تبدأ الحبيبات الإفرازية في التحرك نحو سطح الخلية ، وتتلامس مع غشاء البلازما ، الذي تندمج به أغشيةها الخاصة ، وتكون محتويات الحبيبات خارج الخلية. من الناحية الشكلية ، تسمى هذه العملية البثق ، أو الإخراج (طرد ، خروج الخلايا) وتشبه الالتقام الخلوي ، فقط مع التسلسل العكسي للمراحل.
يمكن أن يزداد حجم مجمع جولجي بشكل كبير في الخلايا التي تؤدي بنشاط الوظيفة الإفرازية ، والتي عادة ما تكون مصحوبة بتطور EPS ، وفي حالة تخليق البروتين ، النواة.
أثناء الانقسام الخلوي ، يتفكك مجمع جولجي إلى صهاريج فردية (ديكتوسومات) و / أو حويصلات ، والتي يتم توزيعها بين خليتين مقسمتين ، وفي نهاية الطور النهائي ، يتم استعادة السلامة الهيكلية للعضية. خارج الانقسام ، هناك تجديد مستمر لجهاز الغشاء بسبب الحويصلات المهاجرة من EPS والصهاريج البعيدة للديكتوسوم بسبب المقصورات القريبة.
إذا وجدت خطأً ، فيرجى تحديد جزء من النص والنقر السيطرة + أدخل.
وزارة التربية والتعليم في جمهورية بيلاروسيا
مؤسسة تعليمية
"الجامعة البيئية الحكومية الدولية
سميت باسم أ. د. ساخاروف »
كلية طب البيئة
جهاز جولجي: الهيكل والوظائف.
طلاب السنة الرابعة
MBD ، غرام. رقم 92062-1
كيسلياتشينكو إيكاترينا
مينسك 2012
مقدمة …………………………………………………………………………… ..… 3
1. هيكل جهاز جولجي ………………………………………………………………… 4
2. وظائف جهاز جولجي ………………………………………………………… .10
3. الآلية الجزيئية لعمل ……………………………… 20
الخلاصة ……………………………………………………………………………………… .22
فهرس
مقدمة
تشكل الشبكة الإندوبلازمية وغشاء البلازما وجهاز جولجي نظامًا لغشاء خلية واحدة ، حيث تحدث عمليات تبادل البروتين والدهون بمساعدة نقل الغشاء داخل الخلايا الموجه والمنظم.
تتميز كل عضيات غشائية بتكوين فريد من البروتينات والدهون.
يتكون جهاز جولجي من مجموعة من الأكياس الغشائية المسطحة - صهاريج ، مجمعة في أكوام - ديكتوسومات (حوالي 5-10 صهاريج ، في حقيقيات النوى السفلية> 30).يختلف عدد الديكتوسومات في الخلايا المختلفة من 1 إلى 500.
صهاريج منفصلة للديكتوسومات ذات السماكة المتغيرة - في وسط أغشيتها يتم تجميعها معًا - تجويف 25 نانومتر ، يتم تشكيل الامتدادات على الأطراف - أمبولات لا يكون عرضها ثابتًا. حوالي 50 نانومتر -1 ميكرومتر فقاعات من الأمبولات المتصلة بالصهاريج بواسطة شبكة من الأنابيب.
في الكائنات متعددة الخلايا ، يتكون جهاز جولجي من أكوام من الصهاريج المترابطة في نظام غشاء واحد. جهاز جولجي عبارة عن نصف كروي ، تواجه قاعدته النواة. يتم تمثيل جهاز الخميرة جولجي بواسطة صهاريج مفردة معزولة محاطة بحويصلات صغيرة وشبكة أنبوبية وحويصلات إفرازية وحبيبات. في طفرات الخميرة Sec 7 و Sec يُظهر الشكل 14 بنية تشبه كومة من خزانات خلايا الثدييات.
يتميز مجمع جولجي بقطبية هياكله. كل كومة لها قطبان: قطب قريب (تشكيل ، سطح رابطة الدول المستقلة) وقاصي (ناضج ، عابر للسطح). القطب المترابط هو جانب الغشاء الذي تندمج به الحويصلات. القطب العابر هو جانب الغشاء الذي تنبت منه الحويصلات.
- هيكل جهاز جولجي.
في عام 1898 ، كشف العالم الإيطالي كاميلو جولجي ، باستخدام خصائص ربط المعادن الثقيلة (الأوزميوم والفضة) مع الهياكل الخلوية ، عن تشكيلات شبكية في الخلايا العصبية ، والتي أطلق عليها اسم "جهاز الشبكة الداخلية" (الشكل 1).
أرز. 1. جهاز شبكي داخل الخلايا (جولجي ، 1898)
أتاح التحسين الإضافي لطريقة تلطيخ المعادن (التشريب) إمكانية التحقق من وجود هياكل شبكية (جهاز جولجي) في جميع خلايا أي كائنات حقيقية النواة. عادة ، توجد عناصر جهاز جولجي (AG) بالقرب من النواة ، بالقرب من مركز الخلية (المريكزات). مناطق جهاز جولجي ، التي تم تحديدها بوضوح بواسطة طريقة التشريب ، في بعض الخلايا كان لها شكل شبكات معقدة ، حيث كانت الخلايا متصلة ببعضها البعض أو ظهرت كمناطق مظلمة منفصلة تقع بشكل مستقل عن بعضها البعض (ديكتوسومات) وتبدو مثل قضبان والحبوب والأقراص المقعرة وغيرها (الشكل 2).

أرز. 2. أنواع جهاز جولجي
أ - شبكة في خلايا ظهارة الأمعاء. ب - منتشر في خلايا العقدة الشوكية.
1 - جوهر 2 - جهاز جولجي ؛ 3 - النواة
لا يوجد فرق جوهري بين الشكلين الشبكي والمنتشر لجهاز جولجي ، لأنه غالبًا ما يتم ملاحظة تغيير في أشكال هذا العضوي في نفس الخلايا. غالبًا ما ترتبط عناصر جهاز جولجي بالفجوات ، وهذا ينطبق بشكل خاص على إفراز الخلايا.
يتغير مورفولوجيا AH تبعًا لمراحل إفراز الخلية ، والتي كانت بمثابة الأساس لـ D.N. Nasonov (1924) طرح فرضية أن AG هي عضية تضمن فصل المواد وتراكمها في مجموعة متنوعة من الخلايا.
لفترة طويلة ، لم يكن من الممكن اكتشاف عناصر جهاز جولجي في الخلايا النباتية باستخدام الأساليب الدقيقة التقليدية. ومع ذلك ، مع ظهور المجهر الإلكتروني ، تم اكتشاف عناصر AG في جميع الخلايا النباتية ، حيث توجد على طول محيط الخلية.
يرتبط وصف هيكل جهاز جولجي ارتباطًا وثيقًا بوصف وظائفه البيوكيميائية الرئيسية ، حيث يتم تقسيم هذه الحجرة الخلوية إلى أقسام بشكل أساسي على أساس توطين الإنزيمات الموجودة في قسم أو آخر.
جهاز جولجي هو جزء متخصص من الشبكة الإندوبلازمية ، ويتكون من أكياس غشائية مسطحة مكدسة. يشارك في إفراز الخلية للبروتينات (يحدث فيها تعبئة البروتينات المفرزة في حبيبات) وبالتالي يتم تطويره بشكل خاص في الخلايا التي تؤدي وظيفة إفرازية.
يتكون مجمع جولجي من خمس حجرات وظيفية:
1. الهياكل الأنبوبية الحويصلة المتوسطة (حجرة وسيطة VTC أو ERGIC - ER - Golgi)
2. Cis-cistern (cis ) - الخزانات القريبة من غرفة الطوارئ:
3. الوسيط (وسطي ) خزانات - خزانات مركزية
4. ترانس-تانك (عبر ) - الأبعد عن خزانات الطوارئ.
5. الشبكة الأنبوبية المجاورة للمحول - شبكة جولجي العابرة ( TGN)

تين. 3. خمسة مكونات ومخطط نقل البروتين.
1. دخول البروتينات المركبة ، والبروتينات السكرية الغشائية والإنزيمات الليزوزومية في خزان ER الانتقالي المجاور لـ AG و 2 - خروجها من ER في الحويصلات المجاورة COPI (النقل المتقدم). 3 - إمكانية نقل البضائع من العناقيد الأنبوبية الحويصلية إلى صهريج AG في الحويصلات COPI ؛ 3 * - نقل البضائع من الخزانات السابقة إلى الخزانات اللاحقة ؛ 4 - إمكانية النقل الحويصلي الرجعي للبضائع بين صهاريج AG ؛ 5- عودة البروتينات المقيمة من AG إلىثالثا مع فقاعات يحدها COPI (النقل إلى الوراء) ؛ 6 و 6 * - نقل الإنزيمات الليزوزومية باستخدام حويصلات مبطنة بالكلاذرين ، على التوالي ، إلى مرحلة مبكرة EE ولاحقًا LE اندوسومات. 7 - إفراز منظم للحبيبات الإفرازية ؛ 8 - التضمين التأسيسي لبروتينات الغشاء في غشاء البلازما القمي للـ PM ؛ 9 ، الالتقام الخلوي بوساطة مستقبلات مع حويصلات مغلفة بالكالاثرين ؛ 10 عودة عدد من المستقبلات من الإندوسومات المبكرة إلى غشاء البلازما ؛ 11- نقل الربيطات من EE إلى LE و lysosomes Ly ؛ 12- نقل الربيطات في حويصلات غير كلاذرين.
تختلف هذه الأقسام عن بعضها البعض بواسطة مجموعة من الإنزيمات. في قسم رابطة الدول المستقلة ، يُطلق على الخزان الأول اسم "صهريج الخلاص" ، حيث إنه بمساعدته تعود المستقبلات القادمة من الشبكة الإندوبلازمية الوسيطة مرة أخرى. إنزيم قسم رابطة الدول المستقلة: فوسفوجليكوزيداز (يعلق الفوسفات على الكربوهيدرات - المانوز). يوجد في القسم الإنسي إنزيمان: ماناسيداز (ينشق مانوز) ون - أسيتيل جلوكوزامين ترانسفيراز (يعلق بعض الكربوهيدرات - الجليكوزامين). يوجد في القسم العابر إنزيمات: ببتيداز (ينفذ تحلل البروتين) وترانسفيراز (ينفذ نقل المجموعات الكيميائية).
جهاز جولجي هو عضية متعددة الأشكال بدرجة عالية. في الخلايا من أنواع مختلفة وحتى في مراحل مختلفة من تطور نفس الخلية ، يمكن أن تبدو مختلفة.
الخصائص الرئيسية لمجمع جولجي هي كما يلي:
- وجود كومة من عدة خزانات (عادة 3-8) بالارض ، متجاورة إلى حد ما مع بعضها البعض. دائمًا ما يكون هذا المكدس محاطًا بعدد معين (مهم جدًا في بعض الأحيان) من حويصلات الغشاء. في الخلايا الحيوانية ، تكون كومة واحدة أكثر شيوعًا ، بينما توجد في الخلايا النباتية عادةً عدة ؛ ثم يتم استدعاء كل منهمالديكتوسوم (الشكل 4).يمكن ربط الديكتوسومات الفردية بنظام من الفجوات ، وتشكيل شبكة ثلاثية الأبعاد ؛

أرز. 4. الترتيب التخطيطي لـ AG في الخلية
2) عدم التجانس التركيبي ، المعبر عنه في حقيقة أن الثوابت (مقيم ) الإنزيمات موزعة بشكل غير متجانس في جميع أنحاء العضية ؛
3) القطبية ، أي وجود جانب رابطة الدول المستقلة يواجه الشبكة والنواة الإندوبلازمية ، والجانب العابر الذي يواجه سطح الخلية (وهذا ينطبق بشكل خاص على الخلايا المفرزة) ؛
4) الارتباط مع الأنابيب الدقيقة ومنطقة المريكز. يؤدي تدمير الأنابيب الدقيقة بواسطة عوامل إزالة البلمرة إلى تفتيت جهاز جولجي ، لكن وظائفه لا تتأثر بشكل كبير. لوحظ تجزئة مماثلة في الظروف الطبيعية ، أثناء الانقسام. بعد استعادة نظام الأنابيب الدقيقة ، يتم جمع عناصر جهاز جولجي المنتشرة في جميع أنحاء الخلية (على طول الأنابيب الدقيقة) في منطقة المركز ، ويتم إعادة بناء مجمع جولجي الطبيعي.
مجمع جولجي عبارة عن كومة من الأكياس الغشائية على شكل قرص (صهريج) ، ممتدة إلى حد ما بالقرب من الحواف ونظام حويصلات جولجي المرتبطة بها. في الخلايا النباتية ، تم العثور على عدد من الأكوام المنفصلة (الديكتوسومات) ، وغالبًا ما توجد في الخلايا الحيوانية واحدة كبيرة أو عدة مكدسات متصلة بواسطة أنابيب.
تنضج البروتينات المعدة للإفراز ، وبروتينات الغشاء الغشائي لغشاء البلازما ، وبروتينات الجسيمات الحالة ، وما إلى ذلك في خزانات جهاز جولجي. تتحرك البروتينات الناضجة بالتتابع على طول صهاريج العضية ، حيث يحدث طيها النهائي ، بالإضافة إلى التعديلات - الارتباط بالجليكوزيل والفسفرة.
جهاز جولجي غير متماثل - الخزانات الموجودة بالقرب من نواة الخلية (cis-Golgi) تحتوي على أقل البروتينات نضجًا ، وحويصلات غشائية - حويصلات ، تتبرعم من الشبكة الإندوبلازمية الحبيبية (ER) ، على الأغشية التي يتم تصنيع البروتينات منها بواسطة الريبوسومات ، متصلة بهذه الدبابات بشكل مستمر.
تحتوي الخزانات المختلفة لجهاز جولجي على إنزيمات حفزية مقيمة مختلفة ، وبالتالي ، تحدث عمليات مختلفة بالتتابع مع البروتينات الناضجة فيها. من الواضح أن مثل هذه العملية التدريجية يجب التحكم فيها بطريقة أو بأخرى. في الواقع ، يتم "تمييز" البروتينات الناضجة ببقايا عديد السكاريد الخاصة (مانوز بشكل أساسي) ، والتي تلعب دور "علامة الجودة" على ما يبدو.
ليس من الواضح تمامًا كيف تتحرك البروتينات الناضجة عبر صهاريج جهاز جولجي بينما تظل البروتينات المقيمة مرتبطة بشكل أو بآخر بصهريج واحد.هناك نوعان من الفرضيات غير الحصرية المتبادلة لشرح هذه الآلية:
- وفقًا للأول ، يتم نقل البروتين باستخدام نفس آليات النقل الحويصلي مثل مسار النقل من ER ، ولا يتم تضمين البروتينات المقيمة في الحويصلة الناشئة.
- وفقًا للثاني ، هناك حركة مستمرة (نضوج) للصهاريج نفسها ، وتجميعها من حويصلات في أحد طرفيها وتفكيكها في الطرف الآخر من العضية ، وتتحرك البروتينات المقيمة إلى الوراء (في الاتجاه المعاكس) باستخدام النقل الحويصلي.
في النهاية ، تتبرعم الحويصلات التي تحتوي على بروتينات ناضجة تمامًا من الطرف المقابل للعضية (عبر جولجي).
يحدث في مجمع جولجي:
- O-glycosylation - ترتبط السكريات المعقدة بالبروتينات من خلال ذرة أكسجين.
- الفسفرة هي إضافة حمض الفوسفوريك إلى البروتينات.
- تشكيل الليزوزوم.
- تكوين جدار الخلية (في النباتات).
- المشاركة في النقل الحويصلي (تكوين تيار ثلاثي البروتين):
- نضج ونقل بروتينات غشاء البلازما ؛
- النضج ونقل الأسرار ؛
- نضج ونقل إنزيمات الليزوزوم.
2. وظائف جهاز جولجي.
وظائف جهاز جولجي متنوعة للغاية.وتشمل هذه:
- فصل البروتين في 3 تيارات:
- تدخل البروتينات الليزوزومية - الغليكوزيلاتي (مع المانوز) إلى قسم رابطة الدول المستقلة من مجمع جولجي ، وبعضها مُفسفر ، ويتم تشكيل علامة من الإنزيمات الليزوزومية - مانوز-6-فوسفات. في المستقبل ، لن تخضع هذه البروتينات المفسفرة للتعديل ، ولكنها ستدخل الجسيمات الحالة.
- الإفراز التأسيسي (الإفراز التأسيسي). يتضمن هذا التدفق البروتينات والدهون ، التي تصبح مكونات الجهاز السطحي للخلية ، بما في ذلك الكاليكس ، أو يمكن أن تكون جزءًا من المصفوفة خارج الخلية.
- الإفراز المستحث - البروتينات التي تعمل خارج الخلية ، الجهاز السطحي للخلية ، في البيئة الداخلية للجسم تصل إلى هنا.خاصية الخلايا الإفرازية.
- تكوين إفرازات مخاطية - جليكوزامينوجليكان (عديدات السكاريد المخاطية)
- تشكيل مكونات الكربوهيدرات في الكاليكس - بشكل رئيسي الشحوم السكرية.
- كبريتات الكربوهيدرات ومكونات البروتين من البروتينات السكرية والجليكوليبيدات
- التحلل البروتيني الجزئي للبروتينات - في بعض الأحيان بسبب هذا ، يصبح البروتين غير النشط نشطًا (يتم تحويل البرونسولين إلى الأنسولين).
الوظيفة الإفرازية لجهاز جولجي.
تشارك عناصر غشاء AG في فصل وتراكم المنتجات المصنعة في ER ، وتشارك في إعادة ترتيبها الكيميائي ، ونضجها: هذا هو بشكل أساسي إعادة ترتيب مكونات السكاريد قليلة السكاريد للبروتينات السكرية في تكوين الأسرار القابلة للذوبان في الماء أو في تكوين الأغشية (الشكل 5).

أرز. الشكل 5. مخطط اتصال الشبكة الإندوبلازمية الحبيبية (ER) ، جهاز جولجي (AG) مع تكوين وإطلاق الزيموجين من خلايا البنكرياس الأسينار
1 - منطقة انتقالية بين EPR و AG ؛ 2 - منطقة نضوج الحبيبات الإفرازية. 3 - حبيبات زيموجين مفصولة عن AG ؛ 4 - خروجهم (خروج الخلايا) خارج الخلية
في خزانات AG ، يتم تصنيع السكريات ، وعلاقتها بالبروتينات ، مما يؤدي إلى تكوين البروتينات المخاطية. ولكن الأهم من ذلك ، بمساعدة عناصر جهاز جولجي ، تتم عملية إزالة الأسرار الجاهزة خارج الخلية. بالإضافة إلى ذلك ، AG هو مصدر للجسيمات الخلوية.
تمت دراسة مشاركة AG في عمليات إفراز المنتجات الإفرازية جيدًا على مثال الخلايا الخارجية للبنكرياس. تتميز هذه الخلايا بوجود عدد كبير من الحبيبات الإفرازية (حبيبات الزيموجين) ، وهي حويصلات غشائية مليئة بمحتوى البروتين. تشتمل بروتينات حبيبات الزيموجين على إنزيمات مختلفة: البروتياز والليباز والكربوهيدرات والنوكليازات. أثناء الإفراز ، يتم إخراج محتويات حبيبات الزيموجين هذه من الخلايا إلى تجويف الغدة ، ثم تتدفق إلى تجويف الأمعاء. نظرًا لأن المنتج الرئيسي الذي تفرزه خلايا البنكرياس هو البروتين ، فقد درسنا تسلسل دمج الأحماض الأمينية المشعة في أجزاء مختلفة من الخلية (الشكل 6). للقيام بذلك ، تم حقن الحيوانات بحمض أميني يسمى التريتيوم (3H-leucine) وتمت مراقبة توطين الملصق في الوقت المناسب باستخدام التصوير الإشعاعي الذاتي المجهري الإلكتروني. اتضح أنه بعد فترة قصيرة من الزمن (3-5 دقائق) ، تم توطين الملصق فقط في المناطق القاعدية للخلايا الغنية بـ ER الحبيبية. منذ أن تم تضمين الملصق في سلسلة البروتين أثناء تخليق البروتين ، كان من الواضح أن تخليق البروتين لا يحدث إما في منطقة AG أو في حبيبات الزيموجين نفسها ، ولكن يتم تصنيعه حصريًا في الإرغستوبلازم على الريبوسومات. بعد ذلك بقليل (بعد 20-40 دقيقة) ، تم العثور على الملصق ، بالإضافة إلى الإرغاستوبلازم ، في منطقة فجوات AG. لذلك ، بعد التوليف في الإرغاستوبلازم ، تم نقل البروتين إلى منطقة AH. حتى في وقت لاحق (بعد 60 دقيقة) ، تم اكتشاف الملصق بالفعل في منطقة حبيبات الزيموجين. في وقت لاحق ، يمكن رؤية العلامة في تجويف أسيني هذه الغدة. وهكذا ، أصبح من الواضح أن AG هو رابط وسيط بين التوليف الفعلي للبروتين المفرز وإزالته من الخلية. كما تمت دراسة عمليات تخليق البروتين وإفرازه بالتفصيل على الخلايا الأخرى (الغدة الثديية ، الخلايا الكأسية للأمعاء ، الغدة الدرقية ، إلخ). كما تمت دراسة السمات المورفولوجية لهذه العملية. يتم فصل البروتين المُصَدَّر المُصنَّع على الريبوسومات وتجميعه داخل صهاريج ER ، حيث يتم نقله إلى منطقة أغشية AG. هنا ، يتم قطع فجوات صغيرة تحتوي على البروتين المركب من المناطق الملساء من ER وتدخل منطقة الفجوة في الجزء القريب من الديكتوسوم. في هذه المرحلة ، يمكن للفجوات أن تندمج مع بعضها البعض ومع الخزان المسطح للديكتوسوم. بهذه الطريقة ، يتم نقل منتج البروتين بالفعل داخل تجاويف خزانات AG.

أرز. 6. تسلسل علامات الكشف (1-4)ا t 3H- ليسين في تخليق وإفراز البروتين من خلية البنكرياس
ك - الشعيرات الدموية ج - سيتوبلازم الخلية. ف - تجويف الغدة. تظهر الأسهم مسارات ترحيل التسمية
نظرًا لتعديل البروتينات في خزانات جهاز جولجي ، يتم نقلها بواسطة فجوات صغيرة من الصهاريج إلى الصهاريج إلى الجزء البعيد من الديكتوسوم حتى تصل إلى شبكة الغشاء الأنبوبي في المنطقة العابرة للديكتوسوم. في هذه المنطقة ، تنقسم الحويصلات الصغيرة التي تحتوي على منتج ناضج بالفعل. يشبه السطح السيتوبلازمي لهذه الحويصلات سطح الحويصلات الحدودية ، والتي يتم ملاحظتها أثناء كثرة الخلايا في المستقبلات. تندمج الحويصلات الصغيرة المنفصلة مع بعضها البعض ، وتشكل فجوات إفرازية. بعد ذلك ، تبدأ الفجوات الإفرازية في التحرك نحو سطح الخلية ، وتتلامس مع غشاء البلازما الذي تندمج به أغشيتها ، وبالتالي تكون محتويات هذه الفجوات خارج الخلية. من الناحية الشكلية ، تشبه عملية البثق (الطرد) كثرة الخلايا ، فقط مع التسلسل العكسي للمراحل. يطلق عليه خروج الخلايا.
مثل هذا الوصف للأحداث ليس سوى مخطط عام لمشاركة جهاز جولجي في عمليات إفرازية. الأمر معقد بسبب حقيقة أن نفس الخلية يمكن أن تشارك في تخليق العديد من البروتينات المُفرزة ، ويمكنها عزلها عن بعضها البعض وتوجيهها إلى سطح الخلية أو في تكوين الجسيمات الحالة. في جهاز جولجي ، لا يتم فقط "ضخ" المنتجات من تجويف إلى آخر ، ولكن يحدث "نضوج" تدريجيًا أيضًا ، وهو تعديل البروتينات ، والذي ينتهي بـ "فرز" المنتجات المتجه إما إلى الجسيمات ، أو إلى غشاء البلازما ، أو إلى فجوات إفرازية.
تعديل البروتين في جهاز جولجي.
تدخل البروتينات التي تم تصنيعها في ER إلى منطقة رابطة الدول المستقلة لجهاز جولجي بعد الارتباط بالجليكوزيل الأولي وتقليل العديد من بقايا السكاريد هناك. في النهاية ، تحتوي جميع البروتينات هناك على نفس سلاسل السكاريد قليلة السكاريد ، التي تتكون من جزيئينن - أسيتيل جلوكوزامين وستة جزيئات مانوز (الشكل 7). في الصهاريج ، يبدأ التعديل الثانوي لسلاسل قليل السكاريد وفرزها إلى فئتين. نتيجة لذلك ، يتم فسفرة السكريات قليلة السكاريد الموجودة على إنزيمات التحلل المائي المخصصة للليزوزومات (السكريات السكرية الغنية بالمانوز) ، كما أن السكريات قليلة السكاريد من البروتينات الأخرى الموجهة إلى الحبيبات الإفرازية أو غشاء البلازما تخضع لتحولات معقدة ، مما يؤدي إلى فقدان عدد من السكريات وإضافة الجالاكتوز ،ن - أسيتيل جلوكوزامين وأحماض السياليك.

أرز. 7. مسارات بروتين سكري الارتباط بالجليكوزيل في جهاز جولجي
أ - بروتينات الحبيبات الإفرازية وغشاء البلازما. ب - بروتينات الليزوزوم. رجل مانوز آسيا والمحيط الهادئ - الأسباراجين. Gl - الجلوكوز SA ، حمض السياليك ؛ ها -ن - أسيتيل الجلوكوزامين. غال - الجالاكتوز
هذا يخلق مجمع خاص من قليل السكريات. يتم إجراء مثل هذه التحولات للسكريات القليلة بمساعدة الإنزيمات - glycosyltransferases ، والتي تعد جزءًا من أغشية خزانات جهاز Golgi. نظرًا لأن كل منطقة في الدكتاتوزومات لها مجموعتها الخاصة من إنزيمات الارتباط بالجليكوزيل ، فإن البروتينات السكرية ، كما كانت ، يتم نقلها من حجرة غشائية ("أرضية" في كومة صهاريج الديكتوسوم) إلى أخرى في سباق التتابع ، وفي كل منها تأثيرات محددة من الإنزيمات. لذلك ، في موقع رابطة الدول المستقلة ، يتم فسفرة المانوز في إنزيمات الليزوزومات وتتشكل مجموعة مانوز 6 فوسفات خاصة ، والتي تتميز بجميع الإنزيمات المتحللة بالماء ، والتي تدخل بعد ذلك في الجسيمات الحالة.
في الجزء الأوسط من الدكتاتوزومات ، يحدث ارتباط ثانوي بالجليكوزيل للبروتينات الإفرازية: إزالة إضافية للمانوز والإضافةن - أسيتيل جلوكوزامين. في المنطقة العابرة ، تضاف أحماض الجالاكتوز والسياليك إلى سلسلة السكريات قليلة السكاريد (الشكل 8).

أرز. 8. توطين الإنزيمات أثناء تعديل البروتين في جهاز جولجي (AG)
1 - تخليق البروتين في ER ؛ 2 - فسفرة السكريات الليزوزومية ؛ 3 - انشقاق المانوز. 4 - اتصالن - أسيتيل الجلوكوزامين. 5 - إضافة المانوز. 6 - إضافة حمض السياليك. 7 - فرز البروتينات على المستقبلات في الشبكة العابرة ؛ 8 - الجسيمات الحالة ؛ 9 - فجوة إفرازية. 10 - البلازما
في عدد من الخلايا المتخصصة في جهاز جولجي ، يتم تركيب السكريات المناسبة.
في جهاز جولجي للخلايا النباتية ، يتم تصنيع السكريات المتعددة لمصفوفة جدار الخلية (الهيميسليلوز ، البكتين). بالإضافة إلى ذلك ، تشارك دبايات الخلايا النباتية في تخليق وإفراز المخاط والموسين ، والتي تحتوي أيضًا على السكريات. يحدث تخليق عديد السكاريد الرئيسي لجدران الخلايا النباتية ، السليلوز ، كما ذكرنا سابقًا ، على سطح غشاء البلازما.
في جهاز جولجي للخلايا الحيوانية ، يتم تصنيع سلاسل عديد السكاريد الطويلة غير الممنوحة من الجليكوزامينوجليكان. أحدها ، حمض الهيالورونيك ، وهو جزء من مصفوفة خارج الخلية للنسيج الضام ، يحتوي على عدة آلاف من كتل السكاريد المتكررة. العديد من الجليكوزامينوجليكان مرتبطة تساهميًا بالبروتينات وتشكل البروتيوغليكان (البروتينات المخاطية). يتم تعديل سلاسل السكاريد هذه في جهاز جولجي وترتبط بالبروتينات التي تفرزها الخلايا في شكل بروتيوغليكان. في جهاز جولجي ، تحدث أيضًا كبريتات الجليكوزامينوجليكان وبعض البروتينات.
فرز البروتين في جهاز جولجي.
تمر ثلاثة تدفقات على الأقل من البروتينات التي صنعتها الخلية عبر جهاز جولجي:
- تدفق الإنزيمات المتحللة للماء في حجرة الجسيمات الحالة ؛
- تدفق البروتينات المفرزة التي تتراكم في فجوات إفرازية ولا يتم إطلاقها من الخلية إلا عند تلقي إشارات خاصة ؛
- تيار من البروتينات الإفرازية المفرزة باستمرار.
لذلك ، يجب أن تكون هناك آلية خاصة للفصل المكاني لهذه البروتينات المختلفة ومساراتها.
في منطقتي رابطة الدول المستقلة والوسطى للديكتوزومات ، تتجمع كل هذه البروتينات معًا دون فصل ، ويتم تعديلها بشكل منفصل فقط اعتمادًا على علامات قليل السكاريد الخاصة بها.
يحدث الفصل الفعلي للبروتينات ، وفرزها ، في المقطع العرضي لجهاز جولجي. لم يتم فك تشفير هذه العملية بشكل كامل ، ولكن باستخدام مثال فرز الإنزيمات الليزوزومية ، يمكن للمرء أن يفهم مبدأ اختيار جزيئات بروتينية معينة (الشكل 9).

أرز. 9. فرز هيدروليسات الحمض في جهاز جولجي (AG)
1 - دخول hydrolases من EPR ؛ 2 - الفسفرة. 3 - النقل إلى شبكة AG العابرة ؛ 4 - الارتباط بالمستقبل ؛ 5 - قشرة الكلاذرين 6 - الجسيم الأولي ؛ 7 - الارتباط بجسيم ثانوي ؛ 8 - الانفصال عن المستقبلات. 9 - نزع الفسفرة. 10 - هيدرولاز المنشط ؛ 11- عودة (إعادة تدوير) المستقبلات
من المعروف أن البروتينات الأولية فقط من hydrolases الليزوزومية لها مجموعة قليلة السكريات محددة ، وهي مجموعة المانوز. في الصهاريج ، يتم فسفرة هذه المجموعات ثم ، مع البروتينات الأخرى ، يتم نقلها من الخزان إلى الخزان عبر المنطقة الوسطى إلى المنطقة العابرة. تحتوي أغشية الشبكة العابرة لجهاز جولجي على بروتين مستقبل عبر الغشاء (مستقبل مانوز -6-فوسفات ، أو مستقبل M-6-P) ، والذي يتعرف على مجموعات مانوز الفسفرة من سلسلة إنزيمات الليزوزومات ويرتبط بها. يحدث هذا الارتباط عند قيم الأس الهيدروجيني المحايدة داخل صهاريج الشبكة العابرة. على الأغشية ، تشكل بروتينات مستقبلات M-6-P عناقيد - مجموعات تتركز في مناطق تكوين حويصلات صغيرة مغلفة بالكلاذرين. في الشبكة العابرة لجهاز جولجي ، يحدث انفصالها وتبرعمها ونقلها إلى الجسيمات الداخلية. لذلك ، فإن مستقبلات M-6-P ، كونها بروتينات عبر الغشاء ، ترتبط بالهيدرولازات الليزوزومية ، وتفصلها (تفرزها) عن البروتينات الأخرى (على سبيل المثال ، الإفرازية ، غير الليزوزومية) وتركزها في حويصلات محدودة. بعد انفصالها عن الشبكة العابرة ، تفقد هذه الحويصلات بسرعة "غلاف" الكلاذرين الخاص بها ، وتندمج مع الإندوسومات ، وتنقل إنزيماتها الليزوزومية المرتبطة بمستقبلات الغشاء إلى هذه الفجوة. كما ذكرنا سابقًا ، يحدث تحمض الوسط داخل الجسيمات الداخلية بسبب نشاط حامل البروتون. بدءًا من الرقم الهيدروجيني 6 ، يتم فصل الإنزيمات الليزوزومية عن مستقبلات M-6-P ، وتنشيطها ، والبدء في العمل في تجويف الجسيم الداخلي. تعود أقسام الأغشية ، جنبًا إلى جنب مع مستقبلات M-6-P ، عن طريق إعادة تدوير حويصلات الغشاء إلى الشبكة العابرة لجهاز جولجي.
على الأرجح ، هذا الجزء من البروتينات الذي يتراكم في فجوات إفرازية ويتم إفرازه من الخلية بعد تلقي إشارة (على سبيل المثال ، عصبي أو هرموني) ، يخضع لنفس إجراء الاختيار (الفرز) على المستقبلات العابرة للصهريج في جولجي جهاز. تدخل هذه البروتينات الإفرازية أولاً في فجوات صغيرة ، تُلبس أيضًا في مادة الكلاذرين ، والتي تندمج بعد ذلك مع بعضها البعض. في الفجوات الإفرازية ، غالبًا ما يحدث تجمع للبروتينات المتراكمة في شكل حبيبات إفرازية كثيفة. وهذا يؤدي إلى زيادة تركيز البروتين في هذه الفجوات بنحو 200 مرة مقارنة بتركيزه في جهاز جولجي. بعد ذلك ، يتم إخراج هذه البروتينات ، وهي تتراكم في فجوات إفرازية ، من الخلية عن طريق الإفراز الخلوي بعد أن تتلقى الخلية الإشارة المناسبة.
يأتي الدفق الثالث من الفجوات المرتبط بالإفراز (التأسيسي) المستمر أيضًا من جهاز جولجي. وهكذا ، تفرز الخلايا الليفية عددًا كبيرًا من البروتينات السكرية والميوسين ، والتي تعد جزءًا من المادة الرئيسية للنسيج الضام. تفرز العديد من الخلايا باستمرار بروتينات تعزز ارتباطها بالركائز. هناك تدفق مستمر من الحويصلات الغشائية إلى سطح الخلية ، وتحمل عناصر من الغشاء المخاطي والبروتينات السكرية. لا يخضع تدفق المكونات التي تفرزها الخلية للفرز في نظام مستقبلات جهاز جولجي. تنفصل الفجوات الأولية لهذا التدفق أيضًا عن أغشية جهاز جولجي وترتبط هيكليًا بالفجوات الحدودية التي تحتوي على الكلاذرين (الشكل 10).

أرز. 10. ثلاثة تيارات لنقل البروتين عبر جهاز جولجي (AG)
1 - التدفق الليزوزومي ؛ 2 - تيار إفراز مستمر ؛ 3 - تدفق الإفراز المنظم
في مجمع جولجي ، لا يحدث فقط نقل الحويصلات من ER إلى غشاء البلازما. هناك نقل رجعي للحويصلات. وهكذا ، تنفصل الفجوات عن الجسيمات الحالة الثانوية وتعاد مع بروتينات المستقبل إلى منطقة trans-AG. بالإضافة إلى ذلك ، هناك تدفق للفجوات من المنطقة العابرة إلى منطقة رابطة الدول المستقلة في AG ، وكذلك من منطقة رابطة الدول المستقلة إلى الشبكة الإندوبلازمية. في هذه الحالات ، تُلبس الفجوات ببروتينات COP.أنا -مركب. يُعتقد أن العديد من إنزيمات الارتباط بالجليكوزيل الثانوية وبروتينات المستقبل في الأغشية يتم إرجاعها بهذه الطريقة.
وفرت هذه السمات لسلوك حويصلات النقل الأساس لفرضية وجود نوعين من نقل مكونات AG (الشكل 11).

أرز. 11. نماذج نقل المنتجات في جهاز جولجي (AG)
أ - نموذج مقصورات مستقرة ؛ 6- نموذج نضج خزانات AG.
1 - إفراز البروتينات. 2 - إنزيمات دائمة AG ؛ 3 - الانتقال إلى الإندوسومات ؛ 4 ، 5 - الانتقال إلى غشاء البلازما ؛أنا - مجمع EPR-AG ؛ثانيًا - قسم رابطة الدول المستقلة من AG ؛ثالثا - القسم المتوسط من AG ؛رابعا - Trans-Uchasyuk AG ؛ V - شبكة Trans-AG
وفقًا لأحدهم ، الأقدم ، في AG ، توجد مكونات غشاء مستقرة ، يتم نقل المواد إليها من ER عن طريق فجوات النقل. وفقًا لنموذج بديل ، AG هو مشتق ديناميكي من ER: بعد أن "يتم ربطه" من ER ، تندمج فجوات الغشاء مع بعضها البعض في خزان جديد ، والذي ينتقل بعد ذلك عبر منطقة AG بأكملها وينقسم في النهاية إلى حويصلات النقل. وفقًا لهذا النموذج ، الرجوع إلى الوراء COP I - تعيد الجزيئات بروتينات AG الدائمة إلى الصهاريج الأصغر.
3. آلية عودة البروتينات الجزيئية لمركب جوجي.
يسمى مركب بروتين عصاري خلوي Heptameric COP I (مجمع غشاء جولجي ، قسيم الغلاف) ، بالتزامن مع GTP -بروتين رابط ARF يشكل الشكل 1 مظروفًا بطريقة ، عند ارتباطه بغشاء جولجي ، يُعتقد أنه يعزز خروج الخلايا الغشائية وتفاعلات الانقسام المرتبطة بنقل غشاء جولجي. تحول على COP I في غشاء جولجي يتطلب وجوده ARF 1 ، والذي يعمل على GTP لكل دورة. ARF 1- GTP تنفذ التضمين COP I في أغشية جولجي ، أثناء التحلل المائي GTP يحفز إطلاق سراح يفترض COP I من الغشاء إلى العصارة الخلوية ، مما يجعل من الممكن تضمينه COP I في دورات دورية لتجميع وتفكيك القشرة. في هذا الطريق، ARF 1 يعمل كمفتاح مزدوج للتحكم في التكامل COP I في الأغشية ، وبالتالي تنظيم وظيفتها.
كان يعتقد في الأصل أن الغشاء ملزم ARF 1 ويشارك قسيم الغلاف بشكل غير انتقائي في تكوين فقاعات النقل. افترض هذا النموذج وجود تدفق كبير للمواد المنقولة عبر المسارات الإفرازية وافترض أن بلمرة قسيم الغلاف ، مدفوعة بالدورة GTP مع ARF 1 يوفر طاقة ميكانيكية-كيميائية لتشكيل الفقاعات. نتيجة لدراسات مختلفة أجريت منذ ذلك الحين ، تم تصحيح وجهة النظر هذه. التنشيط ARF 1 له تأثير كبير على تركيبة الفسفوليبيد للغشاء ويحفز دمج الأكتين وبروتينات العصارة الخلوية الأخرى في غشاء جولجي. هذا يعني القدرة ARF 1 لتسهيل عمليات الفرز والالتقام ورسو أغشية مجمع جولجي.
تم العثور على جزء قسيم الغلاف أيضًا ليكون قادرًا على ربط بقايا ليسين في الشكل الطرفي C لبروتينات الغشاء التي توفر النقل الدوري بين Golgi و ER والوظيفة ، كما هو متوقع ، كسلسلة عودة إلى ER. تتفاعل بطريقة مماثلة مع الأجزاء السيتوبلازمية من بروتينات النقل ، COP I يمكنه تجميع المواد المنقولة في حويصلات والتوسط في فرز بروتينات النقل.
فيما يتعلق بآخر هذه الوظائف ، مسألة ما إذا كان الغشاء مرتبطًا COP I المواد المنقولة إلى حويصلات خارجية أو داخلية أو كلا النوعين.
الخميرة لديها وحدات فرعية متحولة COP I وفقًا لمخطط مصمم لاكتشاف المسوخات غير القادرة على الاحتفاظ / إعادة تدوير الجزيئات الموصوفة بمتبقي ليسين مع الحفاظ على بقية الدورة.
نتيجة لذلك ، تم اقتراح أن بروتينات النقل عبر الغشاء مرتبطة بزخارف تحتوي على الديليسين COP I يتوسط النقل العكسي. ومع ذلك ، مزيد من التحليل للأليلات الفرديةالقسم 21 (جاما COP ) أظهر وجود عيوب انتقائية اعتمادًا على نوع المادة المنقولة في النقل المباشر أيضًا. علاوة على ذلك ، يتعرف قسيم الغلاف أيضًا على التسلسلات المتعلقة بالديليسين والديارجينين في شظايا البروتين السيتوبلازمي.ص 24 ، عائلة كبيرة من النواقل المحتملة التي تكثر في Golgi والتي ثبت أنها تشارك في النقل ثنائي الاتجاه. بالنظر إلى هذه البيانات ، بالإضافة إلى البيانات البيوكيميائية والمورفولوجية السابقة التي تؤكد دور قسيم الغلاف في النقل المباشر ، يصبح الاتجاه (أي المباشر أو العكسي) لنقل الحويصلة غير واضح. والاحتمال الإضافي هو أن الوسيطرابطة ARF 1 COPI باستخدام غشاء يمكن أن يعمل على الفصل الجانبي للبروتينات والدهون في مجموعات منفصلة ، والتي يتم نقلها لاحقًا بطريقة مباشرة أو عكسية. تم اقتراح وجود هذه الميزة كنتيجة لملاحظة حظر الارتباط COP I مع غشاء في طفرات مع تثبيط ARF 1 أو عن طريق العلاج بريفيلدين أ (منتدى بواو الاسيوى ) منع التنشيط ARF 1 لا يتداخل في حد ذاته مع نقل الغشاء ، ولكنه يزعزع استقراره ، مما يؤدي إلى عدم انتقائية عودة البروتين إلى ER.
استنتاج.
مجمع جولجي (جهاز) (أو دكتوسوم) هو عضية غشائية للأغراض العامة تحتوي جميع الخلايا (باستثناء خلايا الدم الحمراء والخلايا الكيراتينية في ظهارة التقرن) على كمية 1 أو أكثر (في تخليق الخلايا بنشاط).
في CG ، يتم الحفاظ على التوازن الديناميكي باستمرار بين عدد الأغشية التي "تترك" جنبًا إلى جنب مع الحويصلات المنقسمة ، وعدد الأغشية التي "تأتي" من ER مع المنتج المركب الذي يحتاج إلى تحسين.
مجمع جولجي عبارة عن هيكل متعدد الوظائف. يؤدي وظائف مختلفة:
1. النقل - ثلاث مجموعات من البروتينات تمر عبر AG: بروتينات الغشاء المحيط بالبروتينات ، والبروتينات المعدة للتصدير من الخلية ، والإنزيمات الليزوزومية.
2. ج الفرز من أجل النقل: الفرز لمزيد من النقل إلى العضيات ، PM ، الإندوسومات ، الحويصلات الإفرازية تحدث في مجمع جولجي العابر.
3. إفراز - إفراز المنتجات المركبة في الخلية.
3. الارتباط بالجليكوزيل للبروتينات والدهون: تزيل الجليكوزيدات بقايا السكر - إزالة الجليكوزيل ، وتربط ناقلات الجليكوزيل السكريات مرة أخرى إلى سلسلة الكربوهيدرات الرئيسية - الارتباط بالجليكوزيل. يحدث الارتباط بالجليكوزيل في سلاسل قليلة السكاريد للبروتينات والدهون ، وكبريتات عدد من السكريات والقصدير ، وكذلك تنشيط سلائف هرمونات عديد الببتيد والببتيدات العصبية.
4. توليف السكريات - يتم تكوين العديد من السكريات في AG ، بما في ذلك البكتين والهيميسليلوز ، والتي تشكل جدران الخلايا في النباتات ومعظم الجليكوزامينوجليكان ، والتي تشكل المصفوفة خارج الخلية في الحيوانات
5. الكبريت - معظم السكريات المضافة إلى لب البروتين للبروتيوجليكان تكون كبريتات
6. إضافة مانوز-6-فوسفات: M-6-ص تمت إضافته كإشارة توجيهية إلى الإنزيمات المخصصة للجسيمات الحالة.
يعد جهاز جولجي جزءًا من جميع الحيوانات تقريبًا (باستثناء خلايا الدم الحمراء في الثدييات) وخلايا النبات.
فهرس.
- أخضر ن . علم الأحياء - م ، 2003
- دي روبرتس إي. نوفينسكي ف ، سايز ف. بيولوجيا الخلية.م ، مير ، 2001
- Zegnbusch P. البيولوجيا الجزيئية والخلوية. م ، مير ، t2004
- Nenitescu K. D. الكيمياء العامة. لكل. من روم. / إد. Ablova A.V - M: Mir ، 1968.
- سفنسون ك. ، ويبستر بي كيج.م ، مير ، 2000.
- سيدوروف إي. علم الأحياء العام - M. ، 2003
- Solovyov Yu. I. ، تطور المشاكل النظرية الرئيسية للكيمياء ، M. ، 1971
- يارجين في. علم الأحياء - م ، 2001